
Geminiviridae (taxid:10811)
VIRION

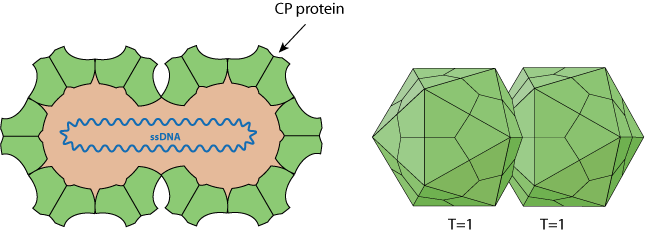
Non-enveloped, about 38 nm in length and 22 nm in diameter (for MSV), twinned (geminate) incomplete T=1 icosahedral symmetry capsid that contains 22 pentameric capsomers made of 110 capsid proteins (CP). Each geminate particle contains only a single circular ssDNA.
GENOME
Mono-, or bipartite, circular, ssDNA genome (+) of about 2.5-3.0 (monopartite) or 4.8-5.6 kb (bipartite). The number of genome segments depends on the genera. 3' terminus has no poly(A) tract. There are coding regions in both the virion (positive) and complementary (negative) sense strands.
The genome is replicated through double-stranded intermediates. The replication (Rep) protein initiates and terminates rolling circle replication, the host DNA polymerase being used for DNA replication itself. There is a potential stem-loop structure in the intergenic region that includes a conserved nona-nucleotide sequence (TAATATTAC) where ssDNA synthesis is initiated.
GENE EXPRESSION
Transcription is bidirectional (c-sense and v-sense). Most of the time, proteins encoded on the c-sense are involved in DNA replication, in regulation of transcription and in interfering with cellular processes, whereas proteins encoded on the v-sense have movement and structural functions.
ENZYMES
REPLICATION
NUCLEAR
- Virus penetrates into the host cell.
- Uncoating, the viral ssDNA genome penetrates into the nucleus.
- The ssDNA is converted into dsDNA with the participation of cellular factors.
- bidirectional dsDNA transcription from the IR promoter produces viral mRNAs and translation of viral proteins.
- Replication is initiated by cleavage of the(+)strand by REP, and occurs by rolling circle producing ssDNA genomes.
- These newly synthesized ssDNA can either
a) be converted to dsDNA and serve as a template for transcription/replication
b) be encapsidated by capsid protein and form virions released by cell lysis.
c) be transported outside the nucleus, to a neighboring cell through plasmodesmata (cell-cell movement) with the help of viral movement proteins.
Host-virus interaction
Cell-cycle modulation
Begomovirus protein Rep and mastrevirus protein RepA are responsible for inhibiting host retinoblastoma protein and inducing transition from the G1 to S phase in preparation for virus replication since the virus targets differentiated non-dividing cells.
 .
.
Suppression of host silencing-mediated antiviral defense
Geminiviruses viral proteins are able to suppress host cell RNA silencing
Begomovirus:
Curtovirus:
Matching UniProtKB/Swiss-Prot entries
(all links/actions below point to uniprot.org website)198 entries grouped by strain
9 entries
Indian cassava mosaic virus (ICMV) reference strain
8 entries
African cassava mosaic virus (isolate West Kenyan 844) (ACMV) (Cassava latent virus (isolate West Kenyan 844)) reference strain
7 entries
Abutilon mosaic virus (isolate West India) (AbMV) reference strain
7 entries
Bean golden yellow mosaic virus (isolate Puerto Rico) (BGYMV) (Bean golden mosaic virus (isolate Puerto Rico)) reference strain
7 entries
Bean golden yellow mosaic virus (isolate Puerto Rico-Japan) (BGYMV) reference strain
7 entries
Beet curly top virus (strain California/Logan) (BCTV) reference strain
7 entries
Cabbage leaf curl virus (isolate Jamaica) (CaLCuV) reference strain
7 entries
Potato yellow mosaic virus (isolate Venezuela) (PYMV) reference strain
7 entries
Squash leaf curl virus (SLCV) reference strain
7 entries
Tomato golden mosaic virus (strain Yellow vein) (TGMV) reference strain
6 entries
Pepper huasteco yellow vein virus (PHYVV) (Pepper huasteco virus) reference strain
5 entries
Tomato leaf curl virus (strain Australia) (ToLCV) reference strain
6 entries
Tomato mottle virus (isolate Florida) (ToMoV) reference strain
6 entries
Tomato pseudo-curly top virus (TPCTV) reference strain
6 entries
Tomato yellow leaf curl China virus (TYLCCNV) reference strain
6 entries
Tomato yellow leaf curl Sardinia virus (TYLCSV) reference strain
6 entries
Tomato yellow leaf curl virus (strain Israel) (TYLCV) reference strain
4 entries
Bean yellow dwarf virus (BeYDV) reference strain
4 entries
Maize streak virus genotype A (isolate South Africa) (MSV) reference strain
4 entries
Maize streak virus genotype C (isolate Set) (MSV) reference strain
4 entries
Maize streak virus genotype D (isolate Raw) (MSV) reference strain
4 entries
Miscanthus streak virus (isolate 91) (MiSV) reference strain
4 entries
Panicum streak virus (isolate Kenya) (PanSV) reference strain
4 entries
Sugarcane streak virus (isolate South Africa) (SSV) (Sugarcane streak virus (isolate Natal)) reference strain
4 entries
Tobacco yellow dwarf virus (strain Australia) (TYDV) reference strain
3 entries
Chloris striate mosaic virus (CSMV) reference strain
8 entries
African cassava mosaic virus (isolate Nigerian) (ACMV) (Cassava latent virus (isolate Nigerian))
8 entries
Mungbean yellow mosaic virus (strain Vigna) (MYMV)
1 entry
Tomato leaf curl virus (strain Australia) (ToLCV)
6 entries
Tomato yellow leaf curl Sardinia virus (isolate Spain-1) (TYLCSV)
6 entries
Tomato yellow leaf curl Sardinia virus (isolate Spain-2) (TYLCSV)
4 entries
Maize streak virus genotype A (isolate Kenya) (MSV)
4 entries
Maize streak virus genotype A (isolate Nigeria) (MSV)
4 entries
Maize streak virus genotype B (isolate Tas) (MSV)
4 entries
Maize streak virus genotype E (isolate Pat) (MSV)
4 entries
Wheat dwarf virus (isolate Sweden) (WDV)
Abutilon mosaic virus (isolate West India) taxid:10816
Ageratum leaf curl virus - [G52] taxid:298650
Ageratum yellow vein Taiwan virus taxid:222441
| Protein | ModelArchive |
| Protein V2 | ma-jd-viral-05659 |
Apple geminivirus PL-2015 taxid:1633752
Axonopus compressus streak virus taxid:1476487
Bean yellow dwarf virus taxid:57119
Begomovirus abutilonbrazilense taxid:1046572
Begomovirus agerati taxid:188333
Begomovirus allamandae taxid:452758
Begomovirus alternantherae taxid:337826
Begomovirus jeskei taxid:932071
Bromus catharticus striate mosaic virus taxid:936005
Capulavirus medicagonis taxid:1306546
Chickpea chlorosis Australia virus taxid:1302848
Chickpea chlorosis virus-A taxid:887825
Chickpea chlorotic dwarf virus taxid:463360
Chickpea redleaf virus taxid:887827
Chickpea yellow dwarf virus taxid:1568974
Citrus chlorotic dwarf associated virus taxid:1202142
Cleome golden mosaic virus taxid:858517
| Protein | ModelArchive |
| Capsid protein (Coat protein) | ma-jd-viral-28407 |
| Replication enhancer (REn) | ma-jd-viral-03817 |
| Transcriptional activator protein (TrAP) | ma-jd-viral-53918 |
Cleome leaf crumple virus taxid:666144
Clerodendron golden mosaic virus taxid:390438
Clerodendrum golden mosaic China virus taxid:559878
Coccinia mosaic Tamil Nadu virus taxid:1532882
Corchorus golden mosaic virus taxid:390436
Corchorus yellow spot virus taxid:403897
Corchorus yellow vein mosaic virus taxid:1297645
Corchorus yellow vein virus - [Hoa Binh] taxid:293284
Cotton leaf crumple virus - [Mexico : Sonora] taxid:335400
Cotton leaf curl Burewala virus taxid:620894
| Protein | ModelArchive |
| C4 protein | ma-jd-viral-57707 |
| Capsid protein (Coat protein) | ma-jd-viral-28153 |
| Protein V2 | ma-jd-viral-05616 |
| Replication-associated protein (Rep) (EC 3.1.21.-) | ma-jd-viral-20433 |
Cotton leaf curl Shahdadpur virus taxid:674996
Cotton leaf curl virus taxid:53010
Cotton yellow mosaic virus taxid:79236
Crassocephalum yellow vein virus - Jinghong taxid:417288
Croton yellow vein mosaic virus taxid:207697
Dalechampia chlorotic mosaic virus taxid:1227356
Datura leaf distortion virus taxid:1227357
Deinbollia mosaic virus taxid:1812308
Desmodium leaf distortion virus taxid:361731
Digitaria ciliaris striate mosaic virus taxid:1196237
Digitaria didactyla striate mosaic virus taxid:889510
Dolichos yellow mosaic virus taxid:333968
Eclipta yellow vein virus taxid:875324
Emilia yellow vein virus-[Fz1] taxid:498805
Eragrostis curvula streak virus taxid:638358
| Protein | ModelArchive |
| Capsid protein (Coat protein) | ma-jd-viral-27737 |
| Movement protein | ma-jd-viral-67039 |
Eragrostis minor streak virus taxid:1030595
Eragrostis streak virus taxid:496807
| Protein | ModelArchive |
| Capsid protein (Coat protein) | ma-jd-viral-27740 |
| Movement protein | ma-jd-viral-54469 |
| Replication-associated protein (Rep) (EC 3.1.21.-) | ma-jd-viral-20157 |
Erectites yellow mosaic virus taxid:390443
Euphorbia caput-medusae latent virus taxid:1853865
Euphorbia leaf curl virus taxid:270117
| Protein | ModelArchive |
| Protein V2 | ma-jd-viral-05722 |
| Replication enhancer (REn) | ma-jd-viral-03776 |
| Transcriptional activator protein (TrAP) | ma-jd-viral-54071 |
Euphorbia leaf curl virus - [G35] taxid:269060
Euphorbia mosaic virus - A [Mexico:Yucatan:2004] taxid:429564
Euphorbia yellow mosaic virus taxid:598494
French bean severe leaf curl virus taxid:1218727
| Protein | ModelArchive |
| Capsid protein (Coat protein) | ma-jd-viral-27802 |
| Replication-associated protein (Rep) (EC 3.1.21.-) | ma-jd-viral-20421 |
Gossypium darwinii symptomless virus taxid:509531
Grapevine geminivirus A taxid:1906317
| Protein | ModelArchive |
| C4 (HAP/C4) | ma-jd-viral-45282 |
| Pre-CP/V2 | ma-jd-viral-11414 |
| Rep/C1 | ma-jd-viral-20636 |
| Replication enhancer (REn) | ma-jd-viral-03577 |
| TrAP/C2 | ma-jd-viral-53903 |
| V1 | ma-jd-viral-27502 |
Grapevine red blotch virus taxid:1381007
Hemidesmus yellow mosaic virus taxid:1383052
Hollyhock leaf crumple virus-[Cairo] taxid:223279
Maize streak Reunion virus taxid:1182518
Malvastrum yellow vein Changa Manga virus taxid:911574
Melon chlorotic mosaic virus taxid:180399
Mesta yellow vein mosaic virus taxid:360579
Mulberry mosaic dwarf associated virus taxid:1631303
Oat dwarf virus taxid:497863
Okra leaf curl virus-[Cameroon] taxid:627503
Paspalum dilatatum striate mosaic virus taxid:1196236
Paspalum striate mosaic virus taxid:1072672
Peristrophe mosaic virus taxid:1188812
| Protein | ModelArchive |
| Movement protein BC1 (Movement protein BL1) | ma-jd-viral-27305 |
| Nuclear shuttle protein (NSP) | ma-jd-viral-27638 |
Saccharum streak virus taxid:683179
| Protein | ModelArchive |
| Capsid protein (Coat protein) | ma-jd-viral-27747 |
| Movement protein | ma-jd-viral-54468 |
| Replication-associated protein (Rep) (EC 3.1.21.-) | ma-jd-viral-20253 |
Soybean chlorotic blotch virus taxid:761702
Soybean mild mottle virus taxid:761701
Spinach curly top Arizona virus taxid:945225
| Protein | ModelArchive |
| C1 | ma-jd-viral-20440 |
| C2 | ma-jd-viral-20957 |
| Capsid protein (Coat protein) | ma-jd-viral-27724 |
| V2 | ma-jd-viral-33808 |
| V3 | ma-jd-viral-30931 |
Spinach severe curly top virus taxid:873160
Sporobolus striate mosaic virus 1 taxid:1302849
Sporobolus striate mosaic virus 2 taxid:1302850
Squash leaf curl Philippines virus taxid:269277
Sugarcane chlorotic streak virus taxid:1919062
Sugarcane white streak virus taxid:1492296
Sunn hemp leaf distortion virus taxid:579113
| Protein | ModelArchive |
| Replication enhancer (REn) | ma-jd-viral-03537 |
| Transcriptional activator protein (TrAP) | ma-jd-viral-54183 |
Sweet potato leaf curl Canary virus taxid:652718
Sweet potato leaf curl Guangxi virus taxid:1489312
Sweet potato leaf curl Henan virus taxid:1489311
Sweet potato leaf curl Henan4 virus taxid:1605995
| Protein | ModelArchive |
| Protein V2 | ma-jd-viral-05720 |
| Transcriptional activator protein | ma-jd-viral-34556 |
Sweet potato leaf curl Sao Paulo virus taxid:994388
Sweet potato leaf curl South Carolina virus taxid:999288
Sweet potato mosaic virus taxid:1513528
Switchgrass mosaic-associated virus 1 taxid:1571533
Tobacco leaf curl virus taxid:67762
Tomato pseudo-curly top virus taxid:49267
Tomato yellow leaf curl virus taxid:10832
| Protein | ModelArchive |
| AC4 (C4) | ma-jd-viral-45383 |
| Protein V2 | ma-jd-viral-05813 |
| Replication enhancer (REn) | ma-jd-viral-03439 |
| Transcriptional activator protein (TrAP) | ma-jd-viral-54051 |
Tomato yellow leaf curl virus (strain Israel) taxid:66366
| Protein | ModelArchive |
| Protein V2 | ma-jd-viral-05813 |
Tomato yellow leaf curl virus-[Almeria] taxid:220938
Turnip curly top virus taxid:859650
| Protein | ModelArchive |
| Capsid protein (Coat protein) | ma-jd-viral-27729 |
| V2 | ma-jd-viral-36572 |
Turnip leaf roll virus taxid:1766828
Watermelon chlorotic stunt virus taxid:35341
Wheat dwarf India virus taxid:1174526
Wheat dwarf virus taxid:10834
| Protein | ModelArchive |
| Capsid protein (Coat protein) | ma-jd-viral-27739 |
| Movement protein | ma-jd-viral-62886 |
| Replication-associated protein (Rep) (EC 3.1.21.-) | ma-jd-viral-60120 |