
Mastrevirus (taxid:10812)
VIRION

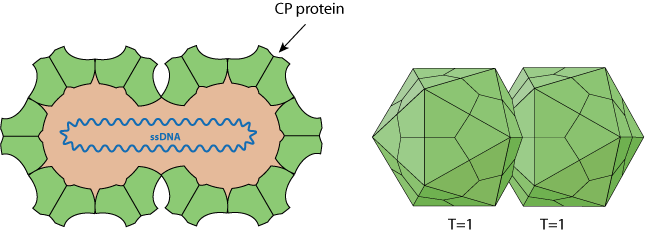
Non-enveloped, about 38 nm in length and 22 nm in diameter (for MSV), twinned (geminate) incomplete T=1 icosahedral symmetry capsid that contains 22 pentameric capsomers made of 110 capsid proteins (CP). Each geminate particle contains only a single circular ssDNA.

GENOME
Monopartite, circular, ssDNA genome (+) genome of about 2.6-2.8 kb. 3' terminus has no poly(A) tract. There are coding regions in both the virion (positive) and complementary (negative) sense strands.
The genome is replicated through double-stranded intermediates. The replication (Rep) protein initiates and terminates rolling circle replication, the host DNA polymerase being used for DNA replication itself. There is a potential stem-loop structure in the long intergenic region (LIR) that includes a conserved nona-nucleotide sequence (TAATATTAC) where ssDNA synthesis is initiated.
The short intergenic region (SIR) contains bidirectional polyadenylation signals.
GENE EXPRESSION
Transcription is bidirectional from the long intergenic region (LIR) which contains two divergent promoters: a V-(virion) and a C-(complementary) sense. The resulting C- and V-transcripts encode four proteins. On the V-sense: the movement and CP. On the C-sense: RepA and Rep (which is expressed as a splice product of RepA and RepB ORFs).
ENZYMES
REPLICATION
NUCLEAR
- Virus penetrates into the host cell.
- Uncoating, the viral ssDNA genome penetrates into the nucleus.
- The ssDNA is converted into dsDNA with the participation of cellular factors.
- bidirectional dsDNA transcription from the IR promoter produces viral mRNAs and translation of viral proteins.
- Replication is initiated by cleavage of the(+)strand by REP, and occurs by rolling circle producing ssDNA genomes.
- These newly synthesized ssDNA can either
a) be converted to dsDNA and serve as a template for transcription/replication
b) be encapsidated by capsid protein and form virions released by cell lysis
c) be transported outside the nucleus, to a neighboring cell through plasmodesmata (cell-cell movement) with the help of viral movement proteins.
Host-virus interaction
Cell-cycle modulation
Mastrevirus protein RepA is responsible for inhibiting host retinoblastoma protein and inducing transition from the G1 to S phase in preparation for virus replication since the virus targets differentiated non-dividing cells.
 .
.
Matching UniProtKB/Swiss-Prot entries
(all links/actions below point to uniprot.org website)55 entries grouped by strain
4 entries
Bean yellow dwarf virus (BeYDV) reference strain
4 entries
Maize streak virus genotype A (isolate South Africa) (MSV) reference strain
4 entries
Maize streak virus genotype C (isolate Set) (MSV) reference strain
4 entries
Maize streak virus genotype D (isolate Raw) (MSV) reference strain
4 entries
Miscanthus streak virus (isolate 91) (MiSV) reference strain
4 entries
Panicum streak virus (isolate Kenya) (PanSV) reference strain
4 entries
Sugarcane streak virus (isolate South Africa) (SSV) (Sugarcane streak virus (isolate Natal)) reference strain
4 entries
Tobacco yellow dwarf virus (strain Australia) (TYDV) reference strain
3 entries
Chloris striate mosaic virus (CSMV) reference strain
4 entries
Maize streak virus genotype A (isolate Kenya) (MSV)
4 entries
Maize streak virus genotype A (isolate Nigeria) (MSV)
4 entries
Maize streak virus genotype B (isolate Tas) (MSV)
4 entries
Maize streak virus genotype E (isolate Pat) (MSV)
4 entries
Wheat dwarf virus (isolate Sweden) (WDV)
Axonopus compressus streak virus taxid:1476487
Bean yellow dwarf virus taxid:57119
Bromus catharticus striate mosaic virus taxid:936005
Chickpea chlorosis Australia virus taxid:1302848
Chickpea chlorosis virus-A taxid:887825
Chickpea chlorotic dwarf virus taxid:463360
Chickpea redleaf virus taxid:887827
Chickpea yellow dwarf virus taxid:1568974
Digitaria ciliaris striate mosaic virus taxid:1196237
Digitaria didactyla striate mosaic virus taxid:889510
Eragrostis minor streak virus taxid:1030595
Eragrostis streak virus taxid:496807
| Protein | ModelArchive |
| Capsid protein (Coat protein) | ma-jd-viral-27740 |
| Movement protein | ma-jd-viral-54469 |
| Replication-associated protein (Rep) (EC 3.1.21.-) | ma-jd-viral-20157 |
Maize streak Reunion virus taxid:1182518
Oat dwarf virus taxid:497863
Paspalum dilatatum striate mosaic virus taxid:1196236
Paspalum striate mosaic virus taxid:1072672
Saccharum streak virus taxid:683179
| Protein | ModelArchive |
| Capsid protein (Coat protein) | ma-jd-viral-27747 |
| Movement protein | ma-jd-viral-54468 |
| Replication-associated protein (Rep) (EC 3.1.21.-) | ma-jd-viral-20253 |
Sporobolus striate mosaic virus 1 taxid:1302849
Sporobolus striate mosaic virus 2 taxid:1302850
Sugarcane chlorotic streak virus taxid:1919062
Sugarcane white streak virus taxid:1492296
Switchgrass mosaic-associated virus 1 taxid:1571533
Wheat dwarf India virus taxid:1174526
Wheat dwarf virus taxid:10834
| Protein | ModelArchive |
| Capsid protein (Coat protein) | ma-jd-viral-27739 |
| Movement protein | ma-jd-viral-62886 |
| Replication-associated protein (Rep) (EC 3.1.21.-) | ma-jd-viral-60120 |