
Mamastrovirus (taxid:249588)
VIRION

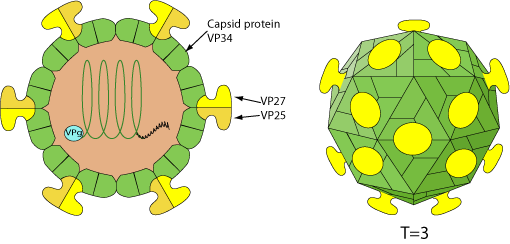
Non-enveloped, spherical, capsid of about 35 nm with T=3 icosahedral symmetry. Surface projections are small and surface appears rough, spikes protruding from the 30 vertices.
VP90 capsid precursor protein undergoes C-terminal cleavages by host caspases to generate VP70 during virus maturation (180 copies of VP70 per particle). Infectious particles are generated by further cleavages of VP70 by extracellular proteases resulting in three structural proteins, VP34, VP27
and VP25  .
.
GENOME

Monopartite, linear ssRNA(+) genome of 6.8-7kb in size. The 5'-terminus is linked to a VPg protein and the 3'-terminus has a poly(A) tract.
GENE EXPRESSION
The virion RNA is infectious and serves as both the genome and viral messenger RNA. The genome contains three overlapping open reading frames (ORF1a, ORF1b, and ORF2). The nonstructural proteins are translated from the genomic RNA as two large polyproteins, nsP1a and nsP1a/1b, through a translational ribosomal frameshifting mechanism. ORF1a and ORF1b encode the viral protease and polymerase respectively (and probably other proteins involved in genome replication). ORF2 is expressed from a subgenomic RNA and encodes the VP90 capsid precursor protein.
ENZYMES
- RNA-dependent RNA polymerase [RdRp]
- VPG-type capping [VPg]
- Polyprotein major protease (Peptidase S39) [Pro]
REPLICATION
CYTOPLASMIC
- Attachement to host receptors probably mediates endocytosis of the virus into the host cell.
- Uncoating, and release of the viral genomic RNA into the cytoplasm.
- Viral RNA is translated into two processed polyproteins to produce replication proteins.
- Replication occurs in viral factories made of membrane vesicles derived from the ER. A dsRNA genome is synthesized from the genomic ssRNA(+).
- The dsRNA genome is transcribed/replicated thereby providing viral mRNAs/new ssRNA(+) genomes.
- Subgenomic RNA translation gives rise to the capsid protein precursor.
- Assembly of new virus particles.
- Non-lytic virus release and maturation of the capsid by proteolytic cleavages.
Host-virus interaction
Adaptive immune response inhibition
Human astrovirus inhibits host complement factors
.
Apoptosis modulation
Host cell apoptosis and host caspases activation seems necessary for maturation of human astrovirus particles .
Matching UniProtKB/Swiss-Prot entries
(all links/actions below point to uniprot.org website)24 entries grouped by strain
3 entries
Human astrovirus-1 (HAstV-1) reference strain
3 entries
Mink astrovirus 1 (MAstV-1) reference strain
3 entries
Ovine astrovirus 1 (OAstV-1) reference strain
1 entry
Human astrovirus VA1 (HAstV-VA1) (Mamastrovirus 9) reference strain
3 entries
Human astrovirus-4 (HAstV-4)
3 entries
Human astrovirus-5 (HAstV-5)
3 entries
Human astrovirus-8 (HAstV-8)
1 entry
Human astrovirus-2 (HAstV-2)
1 entry
Human astrovirus-3 (HAstV-3)
1 entry
Human astrovirus-6 (HAstV-6)
1 entry
Human astrovirus-7 (HAstV-7)
1 entry
Porcine astrovirus 1 (PAstV-1)
Astrovirus wild boar/WBAstV-1/2011/HUN taxid:1137931
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07010 |
| Non-structural polyprotein 1AB | ma-jd-viral-19191 |
| ORF1a | ma-jd-viral-59955 |
Bovine astrovirus taxid:1435485
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07007 |
| NSP1a | ma-jd-viral-59958 |
| Non-structural polyprotein 1AB | ma-jd-viral-40579 |
Bovine astrovirus B170/HK taxid:1027244
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07004 |
| Non-structural polyprotein 1AB | ma-jd-viral-40580 |
| Nsp1a | ma-jd-viral-59961 |
Bovine astrovirus B18/HK taxid:1027245
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07016 |
| Non-structural polyprotein 1AB | ma-jd-viral-40578 |
| Nsp1a | ma-jd-viral-59957 |
Bovine astrovirus B76-2/HK taxid:1027247
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07003 |
| Nsp1a | ma-jd-viral-59945 |
Bovine astrovirus B76/HK taxid:1027248
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07015 |
| Nsp1a | ma-jd-viral-59947 |
Bovine astrovirus CH13 taxid:1522059
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07017 |
| ORF1A nonstructural polyprotein | ma-jd-viral-59964 |
Dromedary astrovirus taxid:1662272
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07012 |
| Nonstructural protein | ma-jd-viral-29454 |
| ORF1a | ma-jd-viral-59963 |
Feline astrovirus 2 taxid:1389204
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-06991 |
| Nsp1a | ma-jd-viral-59940 |
Feline astrovirus D1 taxid:1538452
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07023 |
| Non-structural polyprotein 1AB | ma-jd-viral-19187 |
| ORF1a | ma-jd-viral-59953 |
Human astrovirus BF34 taxid:1518575
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07022 |
| ORF1a | ma-jd-viral-59941 |
Human astrovirus VA1 taxid:645687
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07021 |
| Non-structural protein 1a | ma-jd-viral-59966 |
Ovine astrovirus 1 taxid:1239577
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07018 |
| Non-structural polyprotein 1A | ma-jd-viral-59951 |
Porcine astrovirus 1 taxid:1239567
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-06989 |
Porcine astrovirus 2 taxid:1105378
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07009 |
| ORF1a | ma-jd-viral-59943 |
Porcine astrovirus 3 taxid:1254425
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07020 |
| ORF1a | ma-jd-viral-59949 |
Porcine astrovirus 4 taxid:1105379
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07006 |
| ORF1a | ma-jd-viral-59965 |
Rabbit astrovirus TN/2208/2010 taxid:1161132
| Protein | ModelArchive |
| Capsid polyprotein VP90 | ma-jd-viral-07001 |
| Non-structural polyprotein 1AB | ma-jd-viral-19183 |