
Betaherpesvirinae (taxid:10357)
VIRION

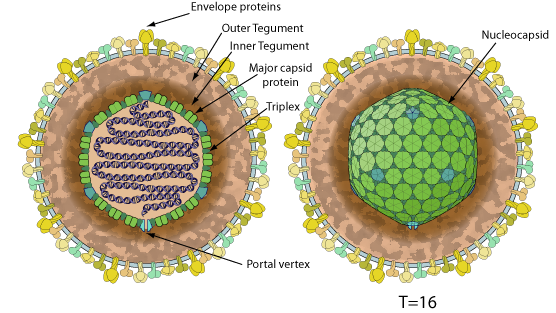
Enveloped, spherical to pleomorphic, 150-200 nm in diameter, T=16 icosahedral symmetry. Capsid consists of 162 capsomers and is surrounded by an amorphous tegument. Glycoproteins complexes are embeded in the lipid envelope.
GENOME
Monopartite, linear, dsDNA genome of 140-240 kb. The genome contains terminal and internal reiterated sequences.
GENE EXPRESSION
Each viral transcript usually encodes a single protein and has a promoter/regulatory sequence, a TATA box, a transcription initiation site, a 5' leader sequence of 30-300 bp (not translated), a 3' nontranslated sequence of 10-30 bp, and a poly A signal. There are many gene overlaps. There are only few spliced genes. Some of the expressed ORFs are antisense to each other. Some ORFs can be accessed from more than one promoter. There are some non-coding genes.
ENZYMES
- DNA-dependent DNA polymerase
- DNA primase
- Tegument deneddylase (Peptidase C76)
- Assemblin (Peptidase S21)
- Kinase
- Helicase
- Ribonucleoside-diphosphate reductase
- Thymidine kinase
- Uracil-DNA glycosylase
REPLICATION
NUCLEAR
Lytic replication:
- Attachment of the viral glycoproteins to host receptors mediates endocytosis of the virus into the host cell.
- Fusion with the plasma membrane to release the core and the tegument proteins into the host cytoplasm.
- The capsid is transported to the nuclear pore where the viral DNA is released into the nucleus.
- Transcription of immediate early genes which promote transcription of early genes and protect the virus against innate host immunity.
- Transcription of early viral mRNA by host polymerase II, encoding proteins involved in replication of the viral DNA.
- A first round of circular genome amplification occurs by bidirectional replication
- Synthesis of linear concatemer copies of viral DNA by rolling circle.
- Transcription of late mRNAs by host polymerase II, encoding structural proteins.
- Assembly of the virus in nuclear viral factories and budding through the inner lamella of the nuclear membrane which has been modified by the insertion of herpes glycoproteins, throughout the Golgi and final release at the plasma membrane.
Latent replication : replication of circular viral episome in tandem with the host cell DNA using the host cell replication machinery.
Host-virus interaction
Adaptive immune response inhibition
Beta-herpesviruses have evolved different strategies to inhibit the host adaptive immune response. For example, HCMV US2 destroys two components of the MHC class II pathway, HLA-DR-alpha and DM-alpha, preventing recognition by CD4+ T cells. Mouse cytomegalovirus (MCMV) uses m152 and m06 encoded proteins to inhibit surface expression of MHC I molecules  .
.
Apoptosis modulation
Apoptosis is very often modulated (and usually inhibited) by beta-herpesviruses. Human cytomegalovirus (HCMV) viral inhibitor of caspase-8-induced apoptosis (from gene UL36) plays a role in the inhibition of apoptosis by interacting with the pro-domain of pro-caspase-8/CASP8 and thus preventing its activation . Murine cytomegalovirus m38.5 protein interacts with host Bax in infected cells and thus inhibits Bax-mediated cell death .
Autophagy modulation
Several beta-herpesviruses are able to inhibit host autophagy process, such as HCMV protein TRS1 via its interaction with host Beclin 1 .
Cell-cycle modulation
The UL24 protein that is present in all herpesvirus subfamilies (alpha, beta and gamma-herpesviruses) induces a cell cycle arrest at G2/M transition through inactivation of the host cyclinB/cdc2 complex .
Innate immune response inhibition
Herpes viruses inhibit the cascade leading to production of interferon-beta by mainly targeting the host IRF3 protein. For example, HCMV targets host IRF3 protein with the viral pp65 protein .
Host splicing inhibition
Modulates the host mRNA expression by exporting unspliced mRNA, thereby inducing alternative splicing .
Matching UniProtKB/Swiss-Prot entries
(all links/actions below point to uniprot.org website)760 entries grouped by protein
6 entries
Alkaline nuclease (EC 3.1.-.-)
1 entry
Protein B1
1 entry
Protein B2
1 entry
Protein B3
1 entry
Protein B4
1 entry
Protein B5
1 entry
Protein B6
1 entry
Protein B7
1 entry
Protein B8
1 entry
Protein B9
2 entries
Putative CC-type chemokine U83
5 entries
Cytoplasmic envelopment protein 1
6 entries
Cytoplasmic envelopment protein 2
6 entries
Cytoplasmic envelopment protein 3
6 entries
Capsid vertex component 1
6 entries
Capsid vertex component 2
1 entry
Chemokine vCXCL1
9 entries
Major DNA-binding protein
12 entries
DNA polymerase catalytic subunit (EC 2.7.7.7)
2 entries
Uncharacterized protein DR1
1 entry
Uncharacterized protein DR2
2 entries
Uncharacterized protein DR3
1 entry
Uncharacterized protein DR4
1 entry
Uncharacterized protein DR5
2 entries
G2/M cell-cycle inhibitor DR6
1 entry
Transactivator protein DR7
1 entry
Uncharacterized protein DR8
7 entries
Deoxyuridine 5'-triphosphate nucleotidohydrolase (dUTPase) (EC 3.6.1.23) (dUTP pyrophosphatase)
2 entries
Early phosphoprotein p84
12 entries
Envelope glycoprotein B (gB)
3 entries
Probable ganciclovir kinase (EC 2.7.1.-)
10 entries
Envelope glycoprotein H (gH)
17 entries
Envelope glycoprotein L (gL)
6 entries
Envelope glycoprotein M (gM)
7 entries
Envelope glycoprotein N
6 entries
120 kDa Glycoprotein O (gO-120K) (Glycoprotein U47)
1 entry
Glycoprotein 105 (gp105)
2 entries
Glycoprotein Q1 (gQ1) (Glycoprotein 105) (gp105) (Glycoprotein Q-80k) (gQ-80k) (HCLF1 protein)
2 entries
Glycoprotein Q2 (gQ2)
5 entries
DNA replication helicase (EC 3.6.4.-)
5 entries
DNA helicase/primase complex-associated protein (HEPA) (Primase-associated factor)
7 entries
mRNA export factor ICP27 homolog
1 entry
Immediate-early protein 1 (IE1) (Protein RF2/RF3/RF4) (pRF2/pRF3/pRF4)
3 entries
Immediate-early protein 2 (IE2)
1 entry
45 kDa immediate-early protein 2 (IE2)
2 entries
Viral interleukin-10 homolog (cmvIL-10) (vIL-10)
2 entries
Protein RL1 (IRL1) (TRL1) (Protein HKLF1)
1 entry
Uncharacterized protein IRL2 (TRL2)
1 entry
Uncharacterized protein IRL3 (TRL3)
1 entry
Uncharacterized protein IRL4 (TRL4)
1 entry
Uncharacterized protein IRL5 (TRL5)
2 entries
Uncharacterized protein IRL6 (TRL6)
1 entry
Uncharacterized protein IRL7 (TRL7)
1 entry
Uncharacterized protein IRL8 (TRL8)
1 entry
Uncharacterized protein IRL9 (TRL9)
2 entries
Protein IRL10 (TRL10)
1 entry
Viral Fc-gamma receptor-like protein IR11 (gp34)
1 entry
Uncharacterized protein IRL12 (TRL12)
1 entry
Uncharacterized protein IRL13 (TRL13)
1 entry
Uncharacterized protein IRL14
2 entries
Protein IRS1
5 entries
Inner tegument protein
1 entry
Uncharacterized protein HKRFX (J1I)
1 entry
Uncharacterized protein J1L
1 entry
Uncharacterized protein J1S
1 entry
Thymidine kinase (EC 2.7.1.21)
8 entries
Large tegument protein deneddylase (EC 3.4.19.12) (EC 3.4.22.-)
1 entry
Glycoprotein M152 (Gp40)
3 entries
Glycoprotein m157
1 entry
Protein M35
6 entries
Major capsid protein (MCP)
6 entries
Nuclear egress protein 1
5 entries
Nuclear egress protein 2
3 entries
Replication origin-binding protein (OBP)
1 entry
Protein ORF A
1 entry
Protein ORF B
1 entry
Protein ORF C
1 entry
Protein ORF D
2 entries
Putative OX-2 membrane glycoprotein homolog (Protein U85)
4 entries
Large structural phosphoprotein (100 kDa phosphoprotein) (pp100) (Major antigenic structural protein)
5 entries
Portal protein
2 entries
Large structural phosphoprotein (150 kDa matrix phosphoprotein) (150 kDa phosphoprotein) (pp150) (Basic phosphoprotein) (BPP) (Phosphoprotein UL32) (Tegument protein UL32)
3 entries
65 kDa phosphoprotein (pp65) (65 kDa matrix phosphoprotein) (Phosphoprotein UL83) (Tegument protein UL83)
2 entries
Protein pp71
4 entries
Phosphoprotein 85 (pp85) (Phosphoprotein UL25)
8 entries
DNA primase (EC 2.7.7.-)
6 entries
Ribonucleoside-diphosphate reductase large subunit-like protein
1 entry
Protein RJ1 (Protein LT1)
1 entry
Membrane glycoprotein RL11
1 entry
Uncharacterized protein RL12
1 entry
Membrane protein RL13
1 entry
Protein RL5A
1 entry
Protein RL8A
1 entry
Protein RL9A
7 entries
Capsid scaffolding protein (Capsid protein P40) (Protease precursor) (pPR)
5 entries
Small capsomere-interacting protein
6 entries
Tegument protein UL51 homolog
1 entry
Tegument protein (67 kDa phosphorylated protein)
1 entry
Protein TRL14
8 entries
Tripartite terminase subunit 1
4 entries
Tripartite terminase subunit 2
6 entries
Tripartite terminase subunit 3 (EC 3.1.-.-) (Terminase large subunit)
2 entries
Protein HHLF1
6 entries
Triplex capsid protein 1
7 entries
Triplex capsid protein 2
3 entries
Protein U10
2 entries
Uncharacterized protein U13
1 entry
Protein UL131A
1 entry
Uncharacterized protein UL147A
1 entry
Protein UL148A
1 entry
Protein UL148B
1 entry
Protein UL148C
1 entry
Protein UL148D
2 entries
Protein U15
1 entry
Uncharacterized protein UL150A
2 entries
Protein U17/U16
3 entries
Protein U17
3 entries
Putative immediate early glycoprotein (Protein U18)
2 entries
Glycoprotein U20
4 entries
U21 glycoprotein
2 entries
Glycoprotein U22
2 entries
Glycoprotein U23
3 entries
U24 protein
1 entry
Uncharacterized protein U24A
1 entry
Protein U54
2 entries
Uncharacterized protein U6
1 entry
Uncharacterized protein U61
1 entry
Protein U62
1 entry
Protein U7
1 entry
Uncharacterized protein U78
4 entries
Protein U79
1 entry
Protein U8
1 entry
Protein U9
1 entry
Protein U90
2 entries
Protein U91
2 entries
Uncharacterized protein U95
2 entries
Glycoprotein UL1
2 entries
Uncharacterized protein UL2
1 entry
Uncharacterized protein UL3
3 entries
Early glycoprotein GP48
2 entries
Protein UL5
2 entries
Uncharacterized protein UL6
2 entries
CEACAM1-like protein UL7
1 entry
Membrane protein UL8
2 entries
Membrane glycoprotein UL9
2 entries
Uncharacterized protein UL10
1 entry
Uncharacterized protein UL101
1 entry
Uncharacterized protein UL106
1 entry
Uncharacterized protein UL107
1 entry
Uncharacterized protein UL108
1 entry
Uncharacterized protein UL109
1 entry
Uncharacterized protein UL110
2 entries
Uncharacterized protein UL116
2 entries
Protein UL117
2 entries
Viral Fc-gamma receptor-like protein UL119 (gp68)
2 entries
Protein UL11
1 entry
Uncharacterized protein UL12
2 entries
Membrane protein UL120
2 entries
Membrane protein UL121
2 entries
Uncharacterized protein UL124
1 entry
Uncharacterized protein UL125
1 entry
Uncharacterized protein UL126
1 entry
Uncharacterized protein UL127
2 entries
Envelope protein UL128
1 entry
Uncharacterized protein UL129
2 entries
Uncharacterized protein UL13
2 entries
Envelope glycoprotein UL130
1 entry
Uncharacterized protein UL131
3 entries
Envelope glycoprotein UL132 (L3)
1 entry
Protein UL133
2 entries
Protein UL135
2 entries
Protein UL136
1 entry
Protein UL138
1 entry
Membrane glycoprotein UL139
2 entries
Uncharacterized protein UL14
1 entry
Protein UL140
1 entry
Protein UL141
1 entry
Membrane glycoprotein UL142
2 entries
Membrane glycoprotein UL144 (TNF alpha-like receptor UL144) (UL144 protein)
1 entry
Protein UL145
2 entries
Putative viral CXC chemokine 2 (vCXCL2)
1 entry
Membrane protein UL148
1 entry
Uncharacterized protein UL150
2 entries
Uncharacterized protein UL15A
2 entries
Protein UL16
3 entries
Protein UL17
2 entries
Glycoprotein UL18
2 entries
Uncharacterized protein UL19
2 entries
Membrane protein UL20
1 entry
Uncharacterized protein UL21
2 entries
Uncharacterized protein UL21A
2 entries
Glycoprotein UL22A
2 entries
Tegument protein UL23
5 entries
Protein UL24 homolog
2 entries
Tegument protein UL26
2 entries
Protein UL27
1 entry
Uncharacterized protein UL28
1 entry
Uncharacterized protein UL29
2 entries
Protein UL29/28
2 entries
Uncharacterized protein UL30
2 entries
Protein UL31
3 entries
Packaging protein UL32
6 entries
G protein-coupled receptor homolog U12
3 entries
Transcriptional regulator UL34
2 entries
Protein UL35
1 entry
Uncharacterized protein UL36
4 entries
Apoptosis inhibitor U19
1 entry
Uncharacterized protein UL39
2 entries
Protein UL40
1 entry
Uncharacterized protein UL41
2 entries
Protein UL41A
3 entries
Protein UL42
2 entries
Tegument protein UL43
5 entries
Protein U33
2 entries
Packaging protein UL32 homolog
1 entry
Uncharacterized protein UL58
1 entry
Uncharacterized protein UL59
1 entry
Uncharacterized protein UL60
1 entry
Uncharacterized protein UL61
1 entry
Uncharacterized protein UL62
1 entry
Uncharacterized protein UL63
1 entry
Uncharacterized protein UL64
1 entry
Uncharacterized protein UL65
1 entry
Uncharacterized protein UL66
1 entry
Uncharacterized protein UL67
1 entry
Uncharacterized protein UL68
1 entry
Uncharacterized protein UL74A
2 entries
Protein UL76
2 entries
Protein UL78
6 entries
Protein U52
1 entry
Uncharacterized protein UL81
3 entries
Protein UL84
6 entries
Protein U58
6 entries
Protein U59
1 entry
Uncharacterized protein UL90
5 entries
Protein U62
6 entries
Protein U63
8 entries
Protein U67
7 entries
Protein U68
5 entries
Phosphotransferase UL97 homolog (EC 2.7.1.-)
5 entries
Uracil-DNA glycosylase (UDG) (EC 3.2.2.27) (UNG)
2 entries
Protein US1
3 entries
Unique short US2 glycoprotein (Protein HQLF2) (gpUS2)
3 entries
Membrane glycoprotein US3
1 entry
Uncharacterized protein US4
2 entries
Uncharacterized protein US5
3 entries
Unique short US6 glycoprotein
2 entries
Membrane glycoprotein US7
2 entries
Membrane glycoprotein US8
2 entries
Membrane glycoprotein US9
2 entries
Membrane glycoprotein US10
3 entries
Membrane glycoprotein US11
2 entries
Uncharacterized protein HVLF6
2 entries
Membrane protein US13
2 entries
Membrane protein US14
2 entries
Membrane protein US15
2 entries
Protein US16
2 entries
Protein US17
3 entries
Transmembrane protein US18
3 entries
Transmembrane protein HWLF4
3 entries
Transmembrane protein HWLF3
2 entries
Membrane protein US21
2 entries
Early nuclear protein HWLF1 (Tegument protein US22)
2 entries
Protein US23
2 entries
Tegument protein US24
1 entry
Uncharacterized protein US25
2 entries
Protein US26
2 entries
Envelope glycoprotein US27
3 entries
G protein-coupled receptor homolog US28 (HHRF3)
2 entries
Uncharacterized protein HHRF4
2 entries
Uncharacterized protein HHRF5
2 entries
Uncharacterized protein HHRF6
2 entries
Protein US32
1 entry
Uncharacterized protein HHLF3
1 entry
Uncharacterized protein US33A
2 entries
Protein US34
2 entries
Uncharacterized protein US34A
1 entry
Uncharacterized protein HHLF2
1 entry
Uncharacterized protein US36
1 entry
30 kDa major early protein
1 entry
Uncharacterized protein U55A
1 entry
Uncharacterized protein U55B
2 entries
G protein-coupled receptor
2 entries
UL37 immediate early glycoprotein
1 entry
Viral inhibitor of caspase-8-induced apoptosis (vICA)
4 entries
Immediate early protein IE1 (IE1) (55 kDa immediate-early protein 1) (IE1p72) (IE72)
4 entries
Viral transcription factor IE2 (IE2) (Protein UL122)
2 entries
30 kDa immediate-early protein 2 (IE2)
2 entries
Protein UL24
5 entries
DNA polymerase processivity factor (Phosphoprotein P41) (PP41) (Polymerase accessory protein) (PAP)
1 entry
Protein U1
1 entry
U10 protein
1 entry
Protein U2
4 entries
Protein U25
5 entries
Protein U26
3 entries
Protein U3
4 entries
Protein U4
1 entry
Protein U5
3 entries
G protein-coupled receptor homolog U51
3 entries
Protein U54
2 entries
Uncharacterized protein U55
1 entry
Protein U7
2 entries
Protein U8
3 entries
Protein U84
2 entries
Protein U9
3 entries
Protein U94
1 entry
Viral Lymphotactin (Viral XCL1) (vSCL1)
1 entry
Uncharacterized protein HHLF4
1 entry
Uncharacterized protein HHRF1
Aotine betaherpesvirus 1 taxid:50290
Caviid betaherpesvirus 2 taxid:33706
Cynomolgus cytomegalovirus taxid:1919083
Elephant endotheliotropic herpesvirus 4 taxid:548914
Elephant endotheliotropic herpesvirus 5 taxid:768738
Elephantid herpesvirus 1 taxid:146015
Elephantid herpesvirus 1 (isolate Asian elephant/Berlin/Kiba/1998) taxid:654902
| Protein | ModelArchive |
| Protein ORF A | ma-jd-viral-36498 |
| Protein ORF B | ma-jd-viral-13328 |
| Protein U4 | ma-jd-viral-13325 |
| Protein U68 | ma-jd-viral-52996 |
| Tegument protein UL51 homolog | ma-jd-viral-35765 |
Guinea pig cytomegalovirus (strain 22122) taxid:103920
| Protein | ModelArchive |
| Envelope glycoprotein B (gB) | ma-jd-viral-31188 |
| Envelope glycoprotein H (gH) | ma-jd-viral-16930 |
| Envelope glycoprotein L (gL) | ma-jd-viral-00343 |
| Protein GP96 | ma-jd-viral-18507 |
Human betaherpesvirus 6A taxid:32603
Human betaherpesvirus 7 taxid:10372
Human cytomegalovirus taxid:10359
Human cytomegalovirus (strain 5508) taxid:69168
| Protein | ModelArchive |
| Envelope glycoprotein L (gL) | ma-jd-viral-00341 |
Human cytomegalovirus (strain AD169) taxid:10360
Human cytomegalovirus (strain Merlin) taxid:295027
Human cytomegalovirus (strain Toledo) taxid:311339
Human cytomegalovirus (strain Towne) taxid:10363
Human herpesvirus 6A (strain GS) taxid:10369
| Protein | ModelArchive |
| Envelope glycoprotein H (gH) | ma-jd-viral-16932 |
Human herpesvirus 6A (strain Uganda-1102) taxid:10370
Human herpesvirus 6B taxid:32604
Human herpesvirus 6B (strain Z29) taxid:36351
Human herpesvirus 7 (strain JI) taxid:57278
Human herpesvirus 7 (strain MUK) taxid:57279
| Protein | ModelArchive |
| Triplex capsid protein 2 | ma-jd-viral-04783 |
Human herpesvirus 7 (strain RK) taxid:262398
| Protein | ModelArchive |
| U21 glycoprotein (gp60) | ma-jd-viral-36538 |