
Orthototiviridae (taxid:3152141)
VIRION

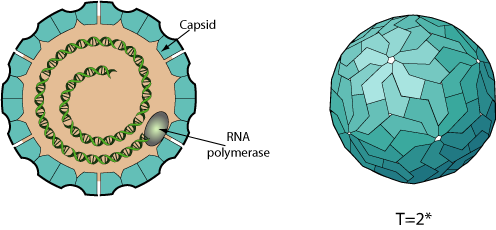
Non enveloped, icosahedral virion composed of a single capsid protein (CP), about 40 nm in diameter. The capsid has a T=2* icosahedral symmetry.

GENOME

Linear dsRNA genome of 4.6-6.7 kb. Contains 2 overlapping ORFs gag and pol, respectively encoding CP and RdRp. Some totiviruses contain a third small 5'-proximal potential ORF.
GENE EXPRESSION
The dsRNA genome is never completely uncoated, to prevent activation of antiviral state by the cell in response to dsRNA. The viral polymerase synthesizes a mRNA, which is translocated to the cell cytoplasm where it is translated.
Translation is initiated on a unique internal ribosome entry site (IRES) element situated at the 5'-UTR.
In most totiviruses, the plus-strand viral transcript is flanked by a 5'untranslated region (5'-UTR) and a 3'-UTR and directs the translation of a major CP (Gag) and a minor fusion protein CP-RdRP (Gag-Pol) via a -1 ribosomal frameshift.
In other totiviruses (e.g. Hv190SV), RDRP is detected only as a separate, non-fused polypeptide and may be expressed through a termination-reinitiation mechanism.
ENZYMES
REPLICATION
CYTOPLASMIC
- Virus either remains intracellular (totivirus) or attachement to host receptors mediates endocytosis of virus into host cell (Giardiavirus).
- Transcription of the dsRNA genome by viral polymerase occurs inside the virion, so that dsRNA is never exposed to the cytoplasm. This plus-strand transcript is used as template for translation.
- (+)RNAs are encapsidated in a sub-viral particle, in which they are transcribed to give RNA (-) molecules with which they become base-paired to produce dsRNA genomes.
- Mature virions are transmitted to new cell during cell division, sporogenesis, cell fusion (totivirus) or are released from the host cell (Giardiavirus).
Matching UniProtKB/Swiss-Prot entries
(all links/actions below point to uniprot.org website)6 entries grouped by strain
2 entries
Saccharomyces cerevisiae virus L-A (ScV-L-A) (ScVL1) reference strain
2 entries
Saccharomyces cerevisiae virus L-BC (ScV-L-BC) (ScVLa) reference strain
1 entry
Ustilago maydis P4 virus (UmV4) (UmV-P4)
1 entry
Ustilago maydis P6 virus (UmV6) (UmV-P6)
Armigeres subalbatus virus SaX06-AK20 taxid:556524
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-55316 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19711 |
Beihai blue swimmer crab virus 3 taxid:1922370
| Protein | ModelArchive |
| Capsid protein | ma-jd-viral-58819 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19940 |
Drosophila melanogaster totivirus SW-2009a taxid:663282
| Protein | ModelArchive |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19707 |
| Structural protein | ma-jd-viral-55315 |
Golden shiner totivirus taxid:1856030
| Protein | ModelArchive |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19714 |
Gremmeniella abietina RNA virus L2 taxid:279539
| Protein | ModelArchive |
| Putative coat protein | ma-jd-viral-15402 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19842 |
Hubei toti-like virus 2 taxid:1923308
| Protein | ModelArchive |
| Major coat protein L-A virus domain-containing protein | ma-jd-viral-21430 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19931 |
Maize-associated totivirus 1 taxid:1729670
| Protein | ModelArchive |
| Capsid protein | ma-jd-viral-32287 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19703 |
Panax notoginseng virus A taxid:1777016
| Protein | ModelArchive |
| Putative coat protein | ma-jd-viral-21434 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19929 |
Penicillium aurantiogriseum totivirus 1 taxid:1755467
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-15415 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19847 |
Red clover powdery mildew-associated totivirus 1 taxid:1714362
| Protein | ModelArchive |
| Capsid protein | ma-jd-viral-21436 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19928 |
Red clover powdery mildew-associated totivirus 2 taxid:1714363
| Protein | ModelArchive |
| Capsid protein | ma-jd-viral-21410 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19945 |
Red clover powdery mildew-associated totivirus 3 taxid:1714364
| Protein | ModelArchive |
| Capsid protein | ma-jd-viral-21438 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19930 |
Red clover powdery mildew-associated totivirus 5 taxid:1714366
| Protein | ModelArchive |
| Capsid protein | ma-jd-viral-61198 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19933 |
Red clover powdery mildew-associated totivirus 6 taxid:1714367
| Protein | ModelArchive |
| Capsid protein | ma-jd-viral-61197 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19936 |
Red clover powdery mildew-associated totivirus 7 taxid:1714368
| Protein | ModelArchive |
| Capsid protein | ma-jd-viral-61199 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19939 |
Saccharomyces cerevisiae virus L-BC taxid:42478
| Protein | ModelArchive |
| Major capsid protein (Gag protein) (Major coat protein) | ma-jd-viral-21444 |
| Probable RNA-directed RNA polymerase (EC 2.7.7.48) (Gag-Pol protein) | ma-jd-viral-19944 |
Saccharomyces kudriavzevii virus L-A1 taxid:1921825
| Protein | ModelArchive |
| Major coat protein | ma-jd-viral-21431 |
Tianjin totivirus taxid:1163537
| Protein | ModelArchive |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19715 |
Wuhan insect virus 26 taxid:1923730
| Protein | ModelArchive |
| Capsid protein | ma-jd-viral-21437 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19935 |
Wuhan insect virus 27 taxid:1923731
| Protein | ModelArchive |
| Major coat protein L-A virus domain-containing protein | ma-jd-viral-21441 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19934 |
Xanthophyllomyces dendrorhous virus L1A taxid:1167690
| Protein | ModelArchive |
| Capsid protein | ma-jd-viral-21440 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19943 |