
Zika virus (taxid:64320)
VIRION

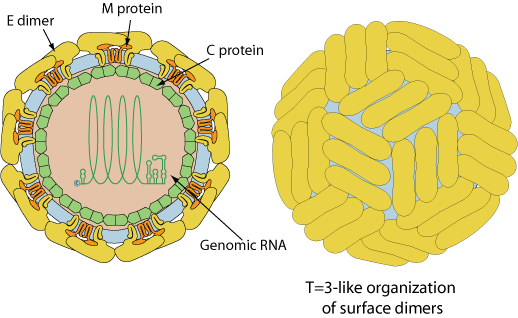
Enveloped, spherical, about 50 nm in diameter. The surface proteins are arranged in an icosahedral-like symmetry.
GENOME

Monopartite, linear, ssRNA(+) genome of of 10,794 bp. The genome 5' end has a methylated nucleotide cap for canonical cellular translation. The 3' terminus is not polyadenylated but forms a loop structure. This secondary structure leads to the formation of a subgenomic flavivirus RNA (sfRNA) through genomic RNA degradation by host XRN1. sfRNA is essential for pathogenicity , and may play a role in inhibiting host RIG-I antiviral activity as shown for Dengue virus
, and may play a role in inhibiting host RIG-I antiviral activity as shown for Dengue virus
GENE EXPRESSION
The virion RNA is infectious and serves as both the genome and the viral messenger RNA. The whole genome is translated in a polyprotein 3,419 aa long, which is processed co- and post-translationally by host and viral proteases. An upstream ORF (uORF) mediates polyprotein translation and would play a role in virus neurotropism .
ENZYMES
- RNA-directed RNA polymerase [NS5]
- Cell-type capping
- Polyprotein major protease (Peptidase S7) [NS3]
REPLICATION
CYTOPLASMIC in mammals,
NUCLEAR in insects?
A. Buckley, E. A. Gould
J. Gen. Virol. August 1988; 69 ( Pt 8): 1913?1920
- Attachement of the viral envelope protein E to host receptors mediates internalization into the host cell by apoptotic mimicry
- Fusion of virus membrane with host endosomal membrane. RNA genome is released into the cytoplasm.
- The positive-sense genomic ssRNA is translated into a polyprotein, which is cleaved into all structural and non structural proteins (to yield the replication proteins).
- Replication takes place at the surface of endoplasmic reticulum in cytoplasmic viral factories. A dsRNA genome is synthesized from the genomic ssRNA(+).
- The dsRNA genome is transcribed/replicated thereby providing viral mRNAs/new ssRNA(+) genomes.
- Virus assembly occurs at the endoplasmic reticulum. The virion buds at the endoplasmic reticulum and is transported to the Golgi apparatus.
- The prM protein is cleaved in the Golgi, thereby maturing the virion which is fusion competent.
- Release of new virions by exocytosis.
Geography
Human cases (source )
S. Ioos, H.-P. Mallet, I. Leparc Goffart, V. Gauthier, T. Cardoso, M. Herida
Med Mal Infect July 2014; 44: 302?307
First report: 1947 in Uganda on a Rhesus monkey
G. W. A. Dick, S. F. Kitchen, A. J. Haddow
Trans R Soc Trop Med Hyg September 1, 1952; 46: 509?520
| Location | Year | Reported Human cases | Estimated Humancases | Seroprevalence |
|---|---|---|---|---|
| 1952 | Uganda | 6.1% | ||
| 1963 | Indonesia | 13% | ||
| 1964 | Uganda | 1 | ||
| 1975 | Nigeria | 2 | ||
| 1977-78 | Indonesia | 17 | ||
| 1999 | Ivory coast | 1 | ||
| 2007 | Micronesia, Yap | 185 | 73% | |
| 2010 | Cambodia | 1 | ||
| 2013 | Australia | 1 (imported) | ||
| 2013 | Indonesia | 1 | ||
| 2013-14 | New-Caledonia | 32 | ||
| 2013-14 | French Polynesia | 8,510 | 29,000 | |
| 2014 | New-Caledonia | 114 | ||
| 2015 | South America | ? | >1.5 million |
Disease
Case fatality rate: <0.001%. Seven death were related to Zika virus infection in Brazil in december 2015, for more than 1 million estimated cases![]() Incubation in Humans: 82 hours
Incubation in Humans: 82 hours
W. G. C Bearcroft
Transactions of the Royal Society of Tropical Medicine and Hygiene September 1956; 50: 438?441
Matching UniProtKB/Swiss-Prot entries
(all links/actions below point to uniprot.org website)8 entries grouped by strain
3 entries
Zika virus (isolate ZIKV/Human/French Polynesia/10087PF/2013) (ZIKV) reference strain
2 entries
Zika virus (ZIKV) reference strain
3 entries