
Victorivirus (taxid:674981)
VIRION

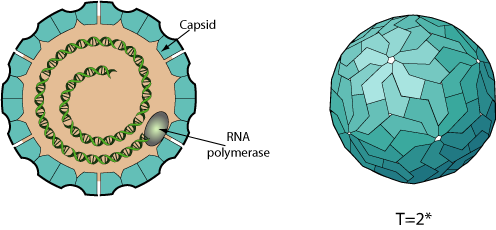
Non enveloped, icosahedral virion composed of a single capsid protein (CP), about 40 nm in diameter. The capsid has a T=2* icosahedral symmetry.
GENOME

Linear dsRNA genome of 4.6-6.7 kb. Contains 2 overlapping ORFs gag and pol, respectively encoding CP and RdRp.
Some isolates contain additional satellite dsRNAs which encode "killer" proteins and are encapsidated separately.
GENE EXPRESSION
The dsRNA genome is never completely uncoated, to prevent activation of antiviral state by the cell in response to dsRNA. The viral polymerase synthesizes a mRNA, which is translocated to the cell cytoplasm where it is translated.
Translation is initiated on a unique internal ribosome entry site (IRES) element situated at the 5'-UTR.
In most totiviruses, the plus-strand viral transcript is flanked by a 5'untranslated region (5'-UTR) and a 3'-UTR and directs the translation of a major CP (Gag) and a minor fusion protein CP-RdRP (Gag-Pol) via a -1 ribosomal frameshift.
In other totiviruses (e.g. Hv190SV), RDRP is detected only as a separate, nonfused polypeptide and may be expressed through a termination-reinitiation mechanism.
ENZYMES
REPLICATION
CYTOPLASMIC
- Virus remains intracellular.
- Transcription of the dsRNA genome by viral polymerase occurs inside the virion, so that dsRNA is never exposed to the cytoplasm. This plus-strand transcript is used as template for translation.
- (+)RNAs are encapsidated in a sub-viral particle, in which they are transcribed to give RNA (-) molecules with which they become base-paired to produce dsRNA genomes.
- Mature virions are transmitted to new cell by cytoplasmic exchange, sporogenesis or hyphal anastomosis.
Matching UniProtKB/Swiss-Prot entries
(all links/actions below point to uniprot.org website)2 entries grouped by strain
2 entries
Helminthosporium victoriae virus-190S (Hv190SV) reference strain
Beauveria bassiana victorivirus NZL/1980 taxid:1485922
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-15413 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19836 |
Botryosphaeria dothidea victorivirus 1 taxid:1547580
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-15419 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19848 |
Chalara elegans RNA Virus 1 taxid:267285
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-15410 |
Fusarium poae victorivirus 1 taxid:1849535
| Protein | ModelArchive |
| Capsid protein | ma-jd-viral-15406 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19831 |
Gremmeniella abietina RNA virus L1 taxid:152217
| Protein | ModelArchive |
| Putative coat protein | ma-jd-viral-15417 |
Magnaporthe oryzae virus 2 taxid:441999
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-15416 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19832 |
Magnaporthe oryzae virus 3 taxid:1661396
| Protein | ModelArchive |
| Putative coat protein | ma-jd-viral-15407 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19827 |
Nigrospora oryzae victorivirus 1 taxid:1765736
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-15411 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19833 |
Penicillium digitatum virus 1 taxid:1833938
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-15409 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19846 |
Phomopsis vexans RNA virus taxid:1580605
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-15420 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19844 |
Rosellinia necatrix victorivirus 1 taxid:1148491
| Protein | ModelArchive |
| Putative capsid protein | ma-jd-viral-15418 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19830 |
Sclerotinia nivalis victorivirus 1 taxid:1859161
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-15408 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19851 |
Tolypocladium cylindrosporum virus 1 taxid:939923
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-15404 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19840 |
Ustilaginoidea virens RNA virus 1 taxid:1312445
| Protein | ModelArchive |
| Putative coat protein | ma-jd-viral-15405 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19843 |
Ustilaginoidea virens RNA virus 3 taxid:1460374
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-15414 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19828 |
Ustilaginoidea virens RNA virus 5 taxid:1756615
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-15412 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19829 |
Ustilaginoidea virens RNA virus L taxid:1561171
| Protein | ModelArchive |
| Coat protein | ma-jd-viral-15424 |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19850 |
Ustilaginoidea virens RNA virus M taxid:1561172
| Protein | ModelArchive |
| RNA-directed RNA polymerase (EC 2.7.7.48) | ma-jd-viral-19739 |