
Passerivirus (taxid:1511802)
VIRION

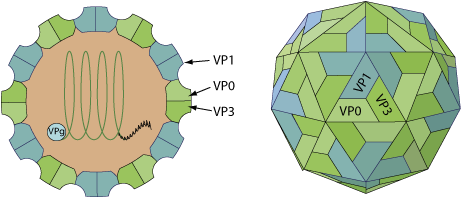
Non-enveloped, spherical, about 30 nm in diameter, pseudo T=3 icosahedral capsid. The capsid consists of a densely-packed icosahedral arrangement of 60 protomers, each consisting of 3 polypeptides, VP0, VP1 and VP3. There is no internal capsid protein.
GENOME

Monopartite, linear, ssRNA(+) genome of about 8 kb, polyadenylated, composed of a single ORF encoding a polyprotein. Viral genomic RNA has a viral protein (VPg) at its 5' end instead of a methylated nucleotide cap structure. The long UTR at the 5' end contains an internal ribosome entry site (IRES). The P1 region encodes the structural polypeptides. The P2 and P3 regions encode the nonstructural proteins associated with replication. Encodes a unique protease 3C. The L protein has been suggested to be involved in RNA replication and encapsidation. The shorter 3' UTR is important in (-)strand synthesis.
GENE EXPRESSION
The virion RNA is infectious and serves as both the genome and viral messenger RNA. The IRES allows direct translation of the polyprotein. The polyprotein is initially processed by the viral protease into various precursor and mature proteins to yield the structural proteins, replicase, VPg, and a number of proteins that modify the host cell, ultimately leading to cell lysis.
ENZYMES
- RNA-dependent RNA polymerase [RdRp]
- VPG-type capping [VPg]
- NTPase-helicase [2C]
- Polyprotein major protease (Peptidase C3) [3Cpro]
REPLICATION
CYTOPLASMIC
- Attachement of the virus to host receptors mediates endocytosis of the virus into the host cell.
- The capsid undergoes a conformational change and possibly opens a pore in the host endosomal membrane and the viral genomic RNA penetrates into the host cell cytoplasm.
- VPg is removed from the viral RNA, which is then translated into a processed polyprotein.
- Replication occurs in viral factories made of membrane vesicles derived from the ER. A dsRNA genome is synthesized from the genomic ssRNA(+).
- The dsRNA genome is transcribed/replicated thereby providing viral mRNAs/new ssRNA(+) genomes.
- New genomic RNA is believed to be packaged into preassembled procapsids.
- Cell lysis and virus release.