
Dianthovirus (taxid:12266)
VIRION

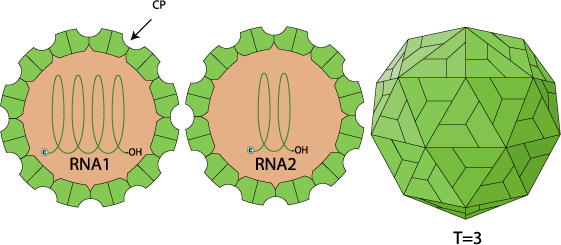
Non-enveloped, spherical, icosahedral, about 28-34 nm in diameter, capsid with a T=3 icosahedral symmetry. 30 hexameric capsomers per nucleocapsid (virion composed of 180 protein subunits).
Genome segments are encapsidated separately into different types of particle

GENOME

Segmented, bipartite, linear ssRNA(+) genome of 3.7-4.0 kb (RNA1) and 1.3-1.4 kb (RNA2). Both RNA1 and RNA2 lack a cap structure and a poly(A)tail.
GENE EXPRESSION
The virion RNA is infectious and serves as both the genome and viral messenger RNA. Two 5'-proximal ORFs are directly translated to produce the viral constituents of the replicase complex. RdRp is translated through ribosomal frameshifting at the end of ORF1. The downstream gene encoding the capsid protein are translated from subgenomic RNAs. The movement protein is translated from RNA-2.
Cap-independent translation is mediated possibly through the interaction between a 3'-cap-independent translation element and a 5'UTR stem-loop.
ENZYMES
REPLICATION
CYTOPLASMIC
- Virus penetrates into the host cell.
- Uncoating, and release of the viral genomic RNA into the cytoplasm.
- The viral RNA is translated to produce the two proteins necessary for RNA synthesis (replication and transcription).
- Replication takes place in cytoplasmic viral factories. A dsRNA genome is synthesized from the genomic ssRNA(+).
- The dsRNA genome is transcribed/replicated thereby providing viral mRNAs/new ssRNA(+) genomes.
- The RdRp recognizes internal subgenomic promoters on the negative-sense RNA to transcribe the 3'co-terminal subgenomic RNAs that will generate the capsid protein. The movement protein is expressed from RNA-2.
- Virus assembly in the cytoplasm.
- Viral movement protein probably mediates virion cell-to-cell transfer.
Matching UniProtKB/Swiss-Prot entries
(all links/actions below point to uniprot.org website)6 entries grouped by protein
2 entries
Capsid protein (Coat protein)
2 entries
Movement protein (MP)
2 entries